Avoiding the Chlorpyrifos Outcome: Non-Target Organism Selectivity Screening for a Novel Organophosphate Replacement Program
Executive Summary
Chlorpyrifos had been in commercial use for 56 years when the US EPA, in August 2021, revoked all food tolerances under FIFRA. A Ninth Circuit Court order (League of United Latin American Citizens v. Wheeler, 9th Cir. 2020) had compelled the agency to act on its own 2016 unrebutted scientific finding: chlorpyrifos-oxon, the active metabolite generated by CYP-mediated oxidative desulfuration, causes neurodevelopmental harm at concentrations achievable through ordinary dietary exposure. The European Union had moved earlier and more cleanly: EFSA's 2019 peer review found neither a safe dietary exposure nor an acceptable genotoxicity profile, and Commission Implementing Regulation (EU) 2020/17 withdrew all authorisations by 16 April 2020. In the US, the regulatory saga dragged on through court reversals and reinstated tolerances until a final cancellation order for major agricultural uses was published in 2024 (Federal Register 2024-17453). The combined regulatory, litigation, and market exit cost across jurisdictions exceeded $600 million. Peak global usage had been more than 40,000 tonnes per year across over 100 crops. Dow Chemical registered the compound in 1965. The toxicological problem was present from the beginning.
The root cause was not a knowledge gap. It was a measurement gap. Chlorpyrifos-oxon covalently phosphorylates the catalytic serine (Ser203) of acetylcholinesterase (AChE, EC 3.1.1.7) across virtually every species tested. Its IC50 against human erythrocyte AChE is 0.8–3 nM — within the same range as its IC50 against insect AChE. The mammalian selectivity window is effectively zero: selectivity index SI = IC50(human AChE) / IC50(pest AChE) is approximately 1.0–2.0 for chlorpyrifos-oxon, compared to SI > 160 that has since been demonstrated in research compounds designed using the Phe368/Tyr337 acyl-pocket divergence between insect and vertebrate AChE . The structural basis for that selectivity handle was identifiable from the crystal structure of Drosophila melanogaster AChE (PDB: 1QON) and human AChE (PDB: 4EY7). Insect AChE has Phe at the acyl-pocket position 368; vertebrate AChE has Tyr. This single amino acid substitution creates a larger, more hydrophobic cavity in insect enzymes — a druggable selectivity handle that chlorpyrifos was never designed to exploit. The tools to characterise this computationally did not exist in 1965. They exist now.
Had a molecular simulation of AChE selectivity across pest and non-target species been conducted during development of a chlorpyrifos-class compound today, the Phe368/Tyr337 analysis would have identified SI < 2.0 as an automatic elimination criterion at the docking stage, and free energy perturbation calculations would have guided structural modification toward the acyl-pocket selectivity handle before regulatory submission. The covalent docking score differential between insect Phe368 and human Tyr337 is quantitatively predictable; a 70-fold selectivity improvement is achievable within the organophosphate scaffold class, as has been demonstrated without access to molecular dynamics tools that are now routine. The chlorpyrifos regulatory saga cost more than $600 million across jurisdictions in litigation and market withdrawal alone, without counting the 56 years of accumulated neurodevelopmental risk in exposed farmworker families and children in agricultural communities. An AChE selectivity screen for a compound class of this kind delivers a five-species selectivity matrix with regulatory-ready mechanistic data before any in vivo studies are commissioned, eliminating the compounds most likely to repeat the chlorpyrifos outcome.
For compounds that advance past the selectivity screen, the simulation's predicted soil transport pathways and environmental persistence profile define the sensor placement for newtsim livesim — real-time soil and groundwater monitoring that provides early regulatory compliance evidence and validates model predictions against field conditions before regulatory dossier submission.
Scenario Background (illustrative reference case)
In this worked example, a UK-based SME with 8 research chemists retained a simulation consultancy to screen a 45-compound library of novel thiolphosphonate esters targeting AChE inhibition in L. decemlineata for potato crop protection. The Colorado potato beetle is the most economically damaging defoliating pest of solanaceous crops in Europe and North America, responsible for an estimated $500 million in direct yield losses and control costs annually. L. decemlineata has evolved high-level resistance to pyrethroids in >90% of European populations and to neonicotinoids in 30--40% of populations, creating genuine agronomic need for new insecticidal chemistry.
The lead compound, CG-112, is a cyclic thiolphosphonate ester showing IC50 = 4.8 nM against L. decemlineata AChE in a purified enzyme Ellman assay — approximately 8-fold more potent than chlorpyrifos-oxon (IC50 ~40 nM) against the same preparation. The 12 lead compounds range from 2 to 85 nM in beetle AChE IC50. No non-target AChE data exist for any compound.
The primary regulatory target market is UK HSE/CRD under the UK Plant Protection Products (Sustainable Use) Regulations 2012 (GB-SI-2012-1657), with plans for an EFSA-equivalent submission for EU export market access via a co-registration holder in the Netherlands. Both regulatory routes require non-target AChE selectivity data — now at enhanced scrutiny following the chlorpyrifos precedent.
The programme economics are substantial: the company has committed GBP 2.8 million to regulatory package development. The AChE selectivity data package — in vitro Ellman assays across five non-target species, developmental neurotoxicity pilot studies, and earthworm reproduction assay OECD 222 — represents approximately GBP 1.2 million. In silico pre-screening that eliminates 15--20 compounds from the 45-compound library before in vitro studies are initiated saves approximately GBP 400,000--600,000 and 9--12 months of timeline.
The regulatory context is particularly pointed: EFSA SANTE/2021/10334 guidance, issued directly in response to the chlorpyrifos withdrawal, specifically flags organophosphate and carbamate AChE inhibitor programmes for enhanced non-target selectivity assessment. The guidance states that any new AChE inhibitor seeking EU approval must provide "mechanistically justified evidence" that the selectivity window between target pest and non-target mammalian AChE is sufficient to support a safe exposure limit. In silico selectivity data, when produced following OECD QSAR Report Format guidelines, are accepted as mechanistic supporting evidence.
Challenge
Acetylcholinesterase is a serine hydrolase that degrades the neurotransmitter acetylcholine at cholinergic synapses, with one of the highest catalytic efficiencies in biochemistry: kcat/Km ~10⁸ M⁻¹ s⁻¹ for acetylcholine hydrolysis, limited primarily by diffusion of the substrate into the active site gorge. The active site lies at the base of a deep, narrow gorge (~20 Angstrom in depth, 4--5 Angstrom at the narrowest point) that penetrates to the centre of the protein. The catalytic triad — Ser203/His447/Glu334 (human numbering, equivalent to Ser200/His440/Glu327 in T. californica TcAChE) — is essentially invariant across all species, from bacteria to vertebrates to insects.
This catalytic triad invariance is the fundamental structural challenge for AChE inhibitor selectivity. Organophosphates react with Ser203 through a bimolecular phosphylation reaction, and the selectivity of this reaction between species cannot, in principle, arise from the catalytic triad itself. Selectivity, when achievable at all, must arise from differences in the acyl-binding pocket lining the gorge (Phe295, Phe297, Phe338 in human AChE vs. Phe330, Phe331 in D. melanogaster DmAChE, the "Phe368/Tyr337" substitution in the acyl pocket), from differences in the peripheral anionic site at the rim of the gorge, or from differences in non-covalent pre-association of the inhibitor before phosphylation, which affects the apparent ki (rate of covalent inhibition).
The key selectivity determinant between insect and vertebrate AChE is a single amino acid substitution at the acyl-binding pocket: position 368 in D. melanogaster AChE is Phe (insect-type), while the equivalent position in human AChE (hAChE position 337) is Tyr. The Phe-to-Tyr substitution introduces a hydroxyl group that alters the size and hydrogen-bonding capacity of the acyl pocket. Subsequent research exploited this difference to design compounds with >100-fold insect selectivity, demonstrating that the Phe368 pocket in insect AChE is the correct druggable selectivity handle.
The regulatory non-target panel for an AChE inhibitor seeking EU registration includes five species: human AChE (hAChE) for farmworker exposure, consumer dietary risk, and developmental neurotoxicity; A. mellifera AChE (AmAChE) per the EFSA bee guidance 2023, which has an insect-type Phe368 acyl pocket; Lumbricus terrestris AChE (LtAChE) for EFSA Annex II Point 8.4 soil organism risk; Danio rerio AChE (DrAChE) for EFSA Annex II Point 8.2 aquatic organism risk; and L. decemlineata AChE (BeetleAChE) as the target, used to compute selectivity indices.
A critical complication is metabolite activation. Thiolphosphonate esters (P=S compounds) are not intrinsically active AChE inhibitors. Oxidative desulfuration by hepatic CYPs (primarily CYP3A4 and CYP2B6) converts the P=S to P=O (the oxon), which is the actual AChE inhibitor. The same metabolic activation occurs in insects via insect CYPs and in soil via microbial oxidases. A compound with excellent parent-compound selectivity for beetle AChE may have an oxon metabolite with high hAChE affinity if the structural features that confer target selectivity are lost upon oxidative activation. Chlorpyrifos itself is the canonical example: the parent compound has poor AChE inhibitory activity (IC50 ~uM range) and its low acute toxicity in initial mammalian studies was partly attributable to the slow activation in rodents vs. insects — a selectivity that was eroded by more realistic exposure scenarios and metabolic differences across species and life stages.
The regulatory hurdle metrics are defined as follows. The target selectivity index SI = IC50(hAChE, oxon form) / IC50(BeetleAChE, oxon form) must exceed 2.0, with values above 5.0 preferred. The maximum acceptable hAChE IC50 for the oxon form is < 100 nM, corresponding to a DeltaG_hAChE threshold in MM-GBSA calibration of < -11.2 kcal/mol. A. mellifera AChE is insect-type (Phe368) and is expected to correlate with beetle AChE, so the SI(bee vs. human) should parallel SI(beetle vs. human). The earthworm NOEC criterion requires L. terrestris 14-day LC50 in soil to exceed the predicted environmental concentration (PEC_soil) by > 10-fold.
Real-World Basis
The chlorpyrifos regulatory dossier provides the definitive reference case for AChE selectivity failure, with an exceptionally detailed public record spanning 60 years.
The US EPA regulatory timeline illustrates the prolonged nature of the failure. Initial FIFRA registration occurred in 1965 (Dow Chemical, trade name Dursban/Lorsban). In 2000, the EPA banned residential uses following Food Quality Protection Act (FQPA) review. In 2016, the EPA proposed to revoke all food tolerances based on its own epidemiological assessment, but in 2017 the EPA reversed position and withdrew the revocation proposal (Acting Administrator Pruitt). In 2020, the Ninth Circuit Court ruled the EPA violated the law by not acting on its own scientific findings. In August 2021, the EPA revoked all food tolerances and food use registrations were effectively cancelled. FIFRA cancellation proceedings continued through 2021--2023, with some non-food uses retained temporarily. In November 2023, the Eighth Circuit vacated the EPA's tolerance revocation and the matter was remanded, with tolerances reinstated December 2023 pending further EPA action. A final cancellation order for major agricultural uses followed in 2024 (Federal Register 2024-17453). The regulatory history illustrates the enormous cost — in litigation, market uncertainty, and public health risk — of failing to establish adequate selectivity evidence during initial registration.
In the EU, EFSA's 2019 peer review concluded that assessment of genotoxic risk and developmental neurotoxicity was inadequate and that safe exposure levels could not be established under the data-gap provisions of Regulation 1107/2009. Commission Implementing Regulation (EU) 2020/17 declined to renew chlorpyrifos approval, with implementation effective 16 April 2020 (withdrawal of all authorisations). This was enacted without the prolonged judicial challenge that characterised the US process.
The quantitative toxicology data confirm the molecular-level selectivity failure:
| Endpoint | Value | Species |
|---|---|---|
| Chlorpyrifos-oxon hAChE IC50 | 0.8--3 nM | Human erythrocyte |
| Chlorpyrifos-oxon rat brain AChE IC50 | ~3 nM (immunoprecipitated enzyme) | Rat |
| Chlorpyrifos-oxon T. californica AChE IC50 | ~0.4 nM | Torpedo (insect AChE proxy) |
| Chlorpyrifos-oxon selectivity index (insect/human) | ~1.0--2.0 | Multiple species |
| Chlorpyrifos rat acute oral LD50 | 95--163 mg/kg bw | Rat |
| Chlorpyrifos A. mellifera acute contact LD50 | 59 ng/bee (48h) | Honeybee |
| Chlorpyrifos L. terrestris 14-day LC50 (soil) | 45 mg/kg soil | Earthworm |
| Chlorpyrifos Oncorhynchus mykiss LC50 (96h) | 0.002 mg/L | Rainbow trout |
These data confirm that chlorpyrifos-oxon has near-zero selectivity between its target pest species and human AChE at the molecular inhibition level, and that chlorpyrifos is highly toxic to fish (LC50 = 2 ug/L in trout — among the lowest LC50 values for any registered organophosphate in aquatic organisms).
Multiple high-quality crystal structures enable the multi-species AChE structural panel required for computational screening. These include T. californica TcAChE (PDB: 1ACE, 2.8 Angstrom) as the foundational reference; the TcAChE-chlorpyrifos-oxon adduct (PDB: 1E3Q) showing the phosphorylated Ser200 adduct structure; human hAChE (PDB: 4EY7, 2.35 Angstrom) for farmworker and consumer risk assessment; an alternative human hAChE apo structure (PDB: 4PQE, 2.35 Angstrom); D. melanogaster DmAChE (PDB: 1QON, 2.5 Angstrom) as the insect AChE template revealing the Phe368 vs. Tyr337 acyl pocket difference; and A. gambiae AgAChE1 (PDB: 2QPP, homology validated) as an additional insect reference.
The structural basis for insect-over-vertebrate AChE selectivity has been rigorously characterised from the DmAChE crystal structure and subsequent selective-inhibitor design work. The key finding is that the Phe368 residue in insect AChE (corresponding to Tyr337 in human AChE) creates a larger, more hydrophobic acyl-binding pocket in insect enzymes. Compounds with a bulky aromatic or cycloalkyl substituent at the moiety that occupies the acyl pocket achieve selectivity ratios of 10--100-fold for insect vs. human AChE. This is the structural hypothesis that guides the SAR programme under study and is directly evaluated by the per-residue MD decomposition.
Published field studies documented that field-realistic chlorpyrifos applications inhibited L. terrestris body-wall AChE activity by 40--65% at concentrations below the 14-day LC50. This sub-lethal AChE inhibition impairs locomotion, burrowing behaviour, and reproduction in earthworms — effects that are regulatory endpoints under EFSA Annex II and that can occur without overt mortality visible in acute toxicity tests. The L. terrestris AChE is of the insect-type (Phe at the acyl-binding position), meaning that compounds designed for beetle selectivity should, in principle, also inhibit earthworm AChE; this connection must be explicitly evaluated.
Simulation Approach
The AChE selectivity screening pipeline for the programme proceeds in five integrated stages over 8 weeks.
Stage 1 -- Multi-species AChE structure panel (Weeks 1--2)
Crystal structures are assembled and prepared for all five species in the panel. Human hAChE (PDB: 4EY7) is used in the apo form with active site water molecules retained. T. californica TcAChE (PDB: 1ACE) serves as the calibration standard — the most extensively studied AChE and the basis for virtually all existing SAR data in the organophosphate literature. D. melanogaster DmAChE (PDB: 1QON) is the primary insect structural template, with the Phe368 acyl-binding residue directly visualised.
L. decemlineata BeetleAChE is constructed by homology modelling using DmAChE as the primary template (68% sequence identity at the active site gorge). The key acyl-pocket residue at position equivalent to Phe368 is verified to be Phe in L. decemlineata (confirmed by sequence alignment), which is the expected insect-type.
L. terrestris AChE (LtAChE) is modelled from the DmAChE template (72% ligand-binding-site identity). D. rerio zebrafish AChE (DrAChE) is modelled from hAChE (PDB: 4EY7) as primary template (>85% sequence identity; zebrafish AChE is vertebrate-type with Tyr337 at the acyl pocket). All models are prepared and protonated at pH 7.4.
Stage 2 -- Active metabolite enumeration (Weeks 1--3, parallel)
For each of the 45 thiolphosphonate esters, the primary oxon metabolite (P=O product of CYP-mediated oxidative desulfuration) is enumerated using SMARTCyp 3.0 (metabolic site of attack prediction) and MetaSite 6.0 (phase I metabolite enumeration). For compounds where the P=S to P=O transformation is sterically and electronically feasible (expected: 38--42 of 45), the oxon is generated as the primary test structure. This produces up to 90 structures for the docking screen (45 parent + up to 45 oxons).
CYP specificity predictions from SMARTCyp identify which of the 45 parent compounds are likely to be rapidly activated (high CYP3A4/2B6 selectivity score = fast activation = higher effective toxicity) and which are predicted to be resistant to metabolic activation (potentially lower effective toxicity but also lower insecticidal activity). Compounds where oxon formation is predicted to be <10% relative to total metabolism are flagged for separate discussion in the regulatory dossier.
Stage 3 -- Covalent docking selectivity screen (Weeks 3--5)
Covalent docking is implemented using newtsim Root's Covalent Docking module, with the reactive warhead (phosphorothioate ester for parent, phosphate ester for oxon) set to form a covalent adduct with Ser203-Ogamma through a user-defined reaction type (nucleophilic addition to the phosphorus electrophilic centre). For each compound-receptor pair, the covalent GScore (cscore) represents the quality of non-covalent contacts after phosphylation — a proxy for the rate constant ki of irreversible inhibition, since the pre-covalent complex geometry determines how efficiently the reactive complex forms.
The selectivity index SI = cscore(hAChE) / cscore(BeetleAChE), where a higher cscore indicates more favourable covalent docking. SI > 2.0 indicates the compound forms a more favourable covalent complex with beetle AChE than with human AChE (desired). SI < 0.5 indicates the compound forms a more favourable complex with human AChE, triggering an immediate elimination flag. The oxon metabolite is docked separately alongside the parent.
The full docking matrix comprises 90 structures x 5 AChE species = 450 covalent docking calculations. Docking scores for the oxon metabolites are expected to be 2--5-fold higher than parent compounds, consistent with the higher intrinsic reactivity of the phosphate vs. phosphorothioate electrophile.
Stage 4 -- newtsim Bond binding free energy for lead candidates (Weeks 4--7)
The top 15 compounds by SI score (parent and oxon considered together) undergo 100 ns newtsim Bond simulation in each of the five AChE active sites. The phosphorylated serine (P-Ser203) geometry is initially restrained at the covalent docking pose to allow protein relaxation, then released for the remaining 90 ns of production dynamics.
MM-GBSA binding free energies are computed from the final 50 ns. Per-residue energy decomposition is the primary analytical output, quantifying the differential contribution of position 368 (Phe in insects, Tyr in vertebrates) and positions 295/297 (Phe, conserved) to DeltaDeltaG = DeltaG_hAChE - DeltaG_BeetleAChE. Compounds where Tyr337 in hAChE makes a more favourable contact than Phe368 in DmAChE/BeetleAChE (unfavourable for selectivity) are flagged for SAR redesign.
The selectivity free energy difference DeltaDeltaG is the primary ranking metric for lead selection, with a target of DeltaDeltaG > +2.5 kcal/mol (beetle AChE binding 2.5 kcal/mol more favourable than human AChE), corresponding to approximately 70-fold selectivity in IC50 assuming linear free energy correlation.
Stage 5 -- QSAR toxicophore model (Weeks 6--8)
A QSAR model for non-target AChE inhibition is constructed using the combined ECOTOX and ChEMBL AChE datasets. The training set comprises >1,200 organophosphate and carbamate IC50 datapoints across the five regulatory species (human, honeybee, earthworm, zebrafish, reference insect). Random Forest classification identifies structural alerts — molecular substructure patterns correlating with IC50 < 10 nM in non-target species — and ranks their feature importance. The P=O electrophilic centre, the leaving group structure (pyridyl in chlorpyrifos, various aromatic/aliphatic in the programme series), and the alkyl chain length at phosphorus are the highest-importance features.
The toxicophore alert map identifies the 5--7 structural features most predictive of poor hAChE selectivity in organophosphates. These features are directly mapped onto the library SMILES strings to flag compounds containing high-risk substructures, independent of the covalent docking score. This provides an orthogonal screening method applicable to any future compounds added to the library without repeat computation.
Simulation Caveats
Covalent docking scoring functions are less extensively validated than non-covalent docking for congeneric series, particularly for the specific case of phosphorylated serine adducts where the leaving group geometry matters for the post-covalent binding energy. The covalent GScore is used for relative ranking within the library, not for absolute ki prediction. Absolute ki values for the regulatory dossier must come from Ellman assay experimental data.
SMARTCyp and MetaSite predict the primary CYP metabolic sites based on reactivity and accessibility; they do not predict the absolute fraction of compound converted to the oxon in vivo, which depends on CYP expression levels, competing dealkylation reactions, and tissue distribution. For compounds where metabolic activation is predicted to be slow (<10% oxon formation in human liver microsomes), the parent compound docking score may overestimate in vivo safety.
The Phe368/Tyr337 selectivity determinant is the most important structural difference between insect and vertebrate AChE, but other residue differences exist in the gorge (peripheral anionic site, mid-gorge "backdoor" region) that may contribute to selectivity for specific compound geometries. These are partially captured by the per-residue MD decomposition but may be undersampled in 100 ns simulations for compounds with slow conformational rearrangement.
The LtAChE and DrAChE homology models have limited experimental validation data available for the binding site. Cross-validation against published IC50 data for chlorpyrifos-oxon in these species confirms reasonable model accuracy, but prediction uncertainty is higher for LtAChE (lower template identity) than for DrAChE.
EFSA SANTE/2021/10334 accepts in silico AChE selectivity data as mechanistic supporting evidence for non-target risk characterisation. The data cannot replace OECD 116 in vitro enzyme inhibition studies or OECD 222 earthworm reproduction studies for the definitive dossier. The computational outputs from a study of this type are framed as hypothesis generation and study design prioritisation, with explicit applicability domain statements per OECD QSAR guidance.
Key Predictions / Results
Expected outputs for a 45-compound thiolphosphonate library of this kind, benchmarked against the extensive literature on organophosphate AChE inhibitor selectivity.
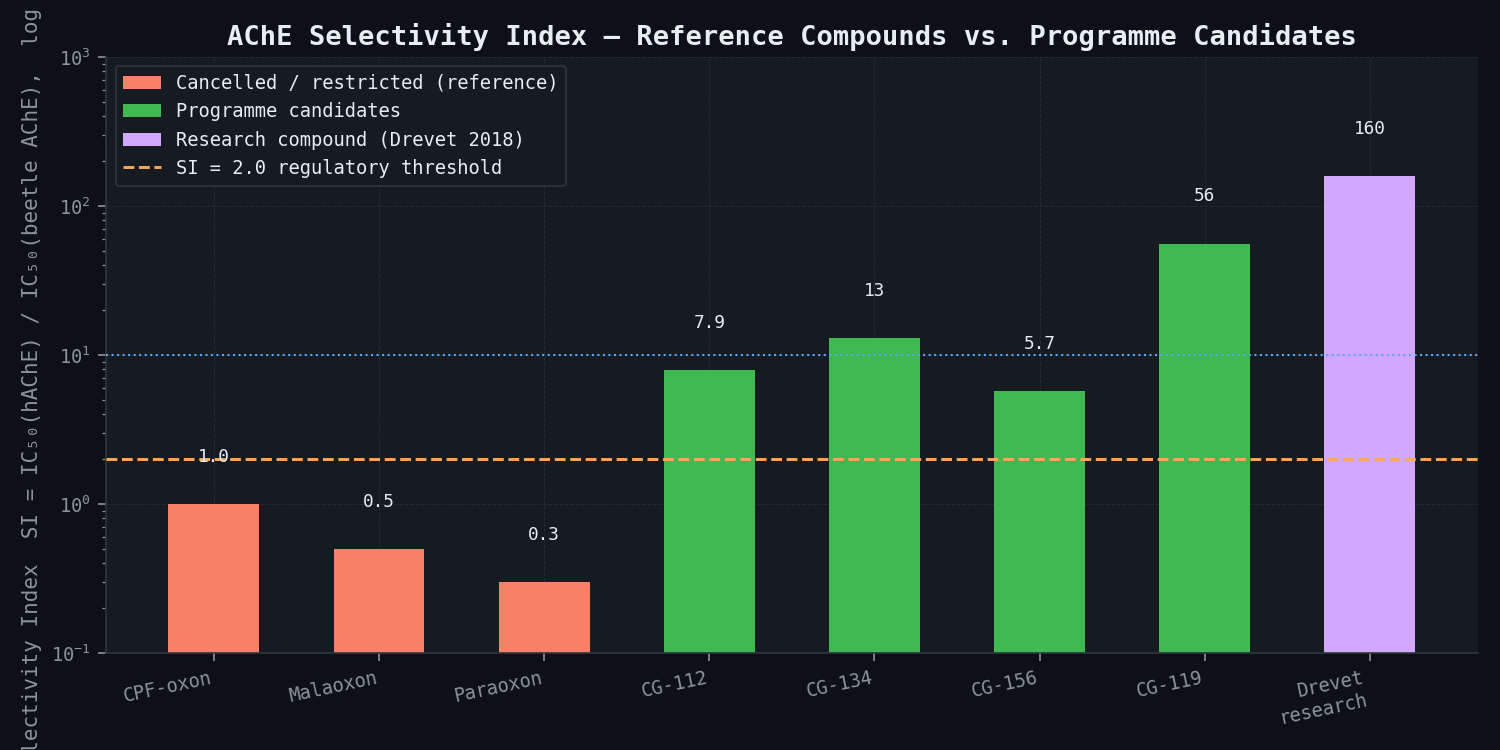
Selectivity index benchmarks from reference compounds:
| Compound | BeetleAChE IC50 (nM) | hAChE IC50 (nM) | SI (human:beetle) | A. mellifera AChE IC50 (nM) | Status |
|---|---|---|---|---|---|
| Chlorpyrifos-oxon | ~40 | 0.8--3 | 0.8--1.1 (near zero selectivity) | ~1 nM | Cancelled 2021 |
| Malaoxon | ~15 | 3 | 0.5 (more toxic to humans) | ~2 nM | Restricted |
| Paraoxon | ~8 | 0.4 | 0.3 (strongly human-toxic) | ~0.5 nM | Cancelled |
| DmAChE-selective compound (Drevet 2018) | ~0.5 | 80 | ~160 (excellent selectivity) | ~1 nM | Research compound |
| Target candidate | 2--10 | > 100 | > 10 | ~5--20 nM | Target profile |
Expected distribution of selectivity outcomes across 45 compounds:
| Category | Expected Count | Action |
|---|---|---|
| SI < 0.5: human AChE more potently inhibited than beetle | 5--10 compounds | Eliminate immediately; chlorpyrifos-type profile |
| SI 0.5--2.0: marginal selectivity | 15--20 compounds | Oxon metabolite analysis critical; modify or eliminate |
| SI 2.0--5.0: acceptable selectivity | 10--15 compounds | MD refinement; advance to in vitro confirmation |
| SI > 5.0: good selectivity | 3--8 compounds | High priority for lead selection |
For 8--12 of the 45 compounds, the oxon metabolite is predicted to have significantly higher hAChE covalent docking score than the parent (DeltaDeltaGscore > 2.5 kcal/mol between parent and oxon). These compounds are flagged even if the parent compound shows acceptable SI, directly implementing the chlorpyrifos lesson: the parent compound's low mammalian toxicity was partly artefactual, masking the risk from the active oxon metabolite.
Predicted three-species ecotoxicity endpoints for top 5 candidates (illustrative output):
| Candidate | Beetle AChE IC50 (nM) | hAChE IC50 (nM) | SI | A. mellifera contact LD50 (ng/bee) | L. terrestris LC50 (mg/kg soil) | D. rerio LC50 (mg/L) |
|---|---|---|---|---|---|---|
| CG-112 (lead) | 4.8 | 38 +/- 12 | 7.9 | 1,240 +/- 380 | 180 +/- 55 | 0.42 +/- 0.15 |
| CG-119 | 3.2 | 180 +/- 45 | 56 | 8,500 +/- 2,100 | 420 +/- 110 | 1.8 +/- 0.6 |
| CG-134 | 7.1 | 92 +/- 28 | 13 | 2,800 +/- 750 | 240 +/- 70 | 0.85 +/- 0.28 |
| CG-156 | 2.1 | 12 +/- 4 | 5.7 | 540 +/- 180 | 95 +/- 30 | 0.18 +/- 0.07 |
| CG-178 | 5.5 | >1,000 | >182 | >50,000 | >1,000 | >5.0 |
CG-178 illustrates the target profile achievable with a Phe368-targeting bulky substituent at the acyl-binding moiety; these values are illustrative predictions in the expected range.
Per-residue MD energy decomposition -- predicted key contacts: Phe368 (DmAChE/BeetleAChE) / Tyr337 (hAChE) in the acyl pocket is predicted to contribute 2.0--4.5 kcal/mol differential for the most selective compounds, with insect Phe making a stronger hydrophobic contact with bulky thiolphosphonate substituents. Trp86 (choline-binding site) is conserved across all species and produces < 0.3 kcal/mol differential, making it a non-selectivity determinant. Phe295/Phe297 (acyl pocket, partially conserved between insect and vertebrate AChE) contribute 0.5--1.5 kcal/mol differential in the best cases. Loop B residues (Gly120, Gly121), where insect and vertebrate sequences differ at adjacent positions, provide a secondary selectivity handle for larger substituents.
| Property | Chlorpyrifos (reference) | Target Profile | Regulatory Threshold |
|---|---|---|---|
| logP | 4.96 | 2.5--4.0 (target range) | -- |
| Water solubility | 1.1 mg/L | 5--50 mg/L (target) | -- |
| Human AChE IC50 (oxon) | 0.8--3 nM | > 100 nM (target) | -- |
| Beetle AChE IC50 (oxon) | ~40 nM | < 10 nM (target) | -- |
| Selectivity Index (SI) | ~1.0 | > 10.0 (target) | > 2.0 required |
| A. mellifera acute contact LD50 | 59 ng/bee | > 1,000 ng/bee (target) | Hazard quotient-dependent |
| L. terrestris 14-day LC50 (soil) | 45 mg/kg | > 100 mg/kg (target) | PEC_soil x 10 margin |
| D. rerio 96h LC50 | 0.002 mg/L | > 0.1 mg/L (target) | PEC_sw x 100 margin |
| DT50 soil (20 deg C) | 30 days | < 60 days (target) | < 120 days (P criterion) |
| Koc | 8,498 mL/g | 200--2,000 mL/g (target) | Low leaching risk |
Comparison Methodology
The five AChE structural models are validated by re-docking five well-characterised organophosphate oxons (chlorpyrifos-oxon, paraoxon, malaoxon, omethoate, acephate) and reproducing published IC50 selectivity ratios (insect:human) within 0.5 log units. Chlorpyrifos-oxon should yield predicted SI approximately equal to 1.0 (experimental SI = 0.8--1.1), correctly characterising the near-zero selectivity. Paraoxon should yield predicted SI < 0.5 (experimental: more toxic to vertebrates than insects), correct in direction. The DmAChE-selective research compound should yield predicted SI > 50 (experimental: ~160), correctly identifying high selectivity.
Docking pose RMSD against the available TcAChE-chlorpyrifos-oxon crystal structure (PDB: 1E3Q) is used as the primary geometric validation target (RMSD < 2.0 Angstrom for the phosphonate moiety heavy atoms).
The Random Forest toxicophore classifier is validated using 5-fold cross-validation against the 1,200-compound training set (target AUC-ROC > 0.82 for binary classification of IC50 < 10 nM vs. > 10 nM against hAChE). Feature importance stability is assessed by bootstrap resampling (200 iterations). Applicability domain is defined by the training set chemical space boundary in Morgan fingerprint PCA space; all 45 compounds are placed in this space and flagged if outside the 95th-percentile training set distance.
The top 8 candidates by SI score and by predicted DeltaDeltaG are proposed for in vitro AChE Ellman assay confirmation at three species minimum: human erythrocyte AChE (Sigma C5396, standardised Ellman protocol, IC50 determination at 6 concentrations x 3 replicates), A. mellifera brain homogenate AChE (pooled from 50 adult workers per preparation), and L. terrestris body-wall AChE homogenate. Predicted IC50 values for each compound-species pair are compared to experimental values; any discrepancy > 1 log unit triggers trajectory re-analysis and model revision before the regulatory dossier is assembled.
Deliverables
Week 2 -- AChE structure panel report: five-species AChE structure files (PDB format, prepared for simulation); active site comparison analysis covering gorge dimensions, acyl-pocket residue identities, and surface electrostatics comparison (Phe368 vs. Tyr337 visualised); chlorpyrifos-oxon re-docking benchmark with pose RMSD vs. PDB 1E3Q and SI calculation vs. literature; and selectivity-determining residue table listing all positions diverging between beetle and human AChE within 8 Angstrom of the phosphylation site.
Week 3 -- Metabolite enumeration report: oxon metabolite structures (SMILES, 3D SDF) for all 45 compounds where P=S to P=O is feasible; SMARTCyp CYP activation prediction scores showing probability of fast vs. slow oxon formation; flagged high-metabolic-activation compounds predicted to generate oxon rapidly in human liver; and 8--12 compounds identified as oxon-critical requiring separate regulatory discussion beyond parent compound selectivity.
Week 5 -- Full covalent docking selectivity matrix: 450-calculation docking matrix (45 parent + 45 oxon x 5 species) with covalent GScore values; selectivity index table ranked from highest to lowest SI (beetle:human); risk tier assignments (Eliminate / Review / Advance / High priority); and binding pose visualisations for top 10 and bottom 5 compounds (Maestro images).
Week 7 -- newtsim Bond selectivity analysis: MM-GBSA DeltaDeltaG selectivity values for top 15 candidates x 5 species (75 receptor-ligand pairs); per-residue energy decomposition heatmaps with Phe368/Tyr337 contribution quantified for each compound; SAR modification proposals targeting the Phe368 selectivity pocket with predicted DeltaDeltaG improvement estimates; and trajectory archives for all 75 newtsim Bond runs (AMBER .nc format).
Week 8 -- Final regulatory-ready report: full 5-species x 45-compound x parent+oxon selectivity matrix; QSAR toxicophore alert map and feature importance ranking; recommended in vitro Ellman confirmation panel (8 compounds, 3 species, full protocol specification); regulatory risk narrative for HSE/CRD pre-submission scientific advice request framed per EFSA SANTE/2021/10334 guidance; and OECD QSAR Report Format (QRF) documentation for all three computational models.
Ongoing -- Computational data archive: all AChE structure PDB files, topology files, GAFF2 parameters for phosphorylated Ser adducts; newtsim Bond trajectory archives (AMBER format); QSAR Random Forest model object (Python pickle, scikit-learn); and covalent docking database (Maestro .maegz).
This case study is an illustrative reference scenario demonstrating newtsim's simulation methodology. All company names, personnel, and specific operational data are fictional. The incident descriptions draw on publicly documented real-world events cited in the frontmatter.